
ALOCASIA BRISBANENSIS
SYNONYMS:
HOMOTYPIC SYNONYMS: Alocasia macrorrhizos var. brisbanensis
HETEROTYPIC SYNONYMS: Caladium macrorrhizon, Colocasia macrorrhizos [‘-rrhiza’], Caladium glycyrrhizon
ACCEPTED INFRASPECIFICS: N/A
DISTRIBUTION: Australia | Queensland & New South Wales (up to Shoalhaven river); New Zealand (introduced)
CLIMATE: Subtropical Humid climate
Humidity is moderate throughout the year, ranging from 60% to 70%
Temperature is varies between the seasons - within the range of 48°F/9°C to 88°F/31°C during the day. Minimum temperatures never dip below 45°F/7°C
Rainy and humid season (October to May) and a dry season between June and October. The average annual rainfall is 1,200 mm
ECOLOGY: Found in tropical, subtropical, and warm temperate rainforest, and wet open sites, predominantly at low altitudes, generally on pleiotrophic soils, along creek lines and moist forest floors
SPECIES DESCRIPTION:
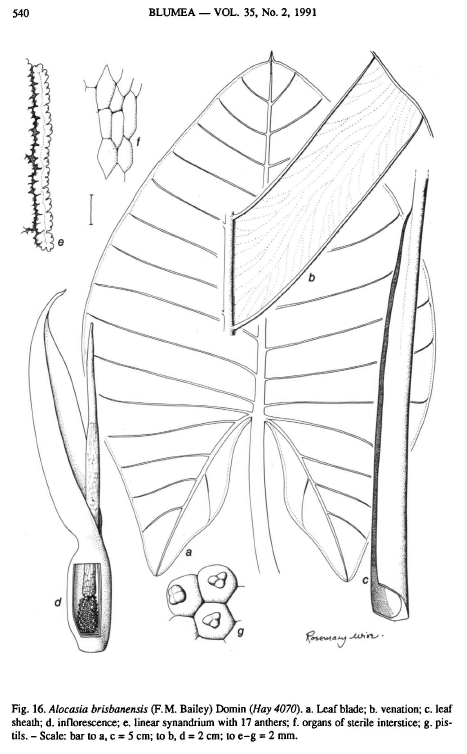
Massive herb; stem decumbent to erect, to c. 13 cm diam., clothed in the brown remains of old leaf bases; leaves several together; petiole to c. 1.1m long, sheathing in the lower c. 2/3rds, eglandular; wings of sheath persistent, straight, sometimes distally reflexed; blade sagittate to somewhat ovato-sagittate, membranous, pale to mid green, with the margin entire; anterior lobe to c. 60 cm long, c. 60 cm wide at base, with the distal part narrowly obtuse and the tip apiculate; anterior costa with c. 11 primary lateral veins on each side diverging at c. 45-60 degrees; primary venation prominent and secondary venation flush on both surfaces; interprimary collective veins present to very ill-defined; glands in the axils of primary veins absent or extremely inconspicuous; posterior lobes c. 30 cm long, ± triangular with well-defined, acute to rectangular, narrowly rounded tips; posterior costae not or hardly naked in the sinus, or the blade very slightly peltate
Shoot growth in Alocasia brisbanensis is sylleptic (new shoots develop simultaneously with inflorescence, without a resting phase).
Petiole sheathing about two-thirds of its length, persistent wings, blade often slightly peltate, interprimary veins present but ill-defined.
Leaf morphology: Posterior lobes have acute to rectangular, narrowly rounded tips. Veins prominent, glands at vein axils either absent or inconspicuous.
INFLORESCENCE:
Inflorescence solitary (in young plants, or first of flowering episode) to paired, on long peduncles about half the length of, to equaling, rarely exceeding the petiole, subtended by a large, persistent, eventually marcescent cataphyll; spathe c. 25 cm long, distinctly constricted between 1/4 and 1 /3rd of the way from the base; the limb opening wide, with the lower part deflected, the upper part arching over the tip of the spadix but not cowl-like, later all deflected, green to yellowish green to white (?), distinctly thick and leathery, marcescent, not deliquescent; spadix sessile, the female zone c. 3 cm long, c. 1.5 cm diameter, with c. 120 pistils; ovary globose, pale green, unilocular, apically incompletely 3-septate; style very short; stigma with 3 or 4 blunt lobes, green; ovules several, hemianatropous; interstice of sterile organs to 4.2 cm long, tapering distally to c. 7 mm diam., often with a short naked zone between it and the female zone; sterile organs numerous and ± globose below, longitudinally confluent and grooved above, green to pale orange; male zone c. 8 cm long, c. 1.5 cm diam., green to greenish yellow; synandria ± linear, longitudinally orientated, 2-36-staminate, 0.2- 2 cm long, c. 2 mm wide with the tops longitudinally grooved; anthers opening by apical pores; appendix subequalling the male zone, tapering to a point, composed of numerous staminodes, green, becoming orange; fruit a several-seeded bright red berry exposed by the longitudinally recurved segments of the dehiscent lower spathe; seeds several, brown to black, c. 3-4 mm diameter, more or less beaked strophiolate.Spathe limb thick, leathery, and marcescent (withers but doesn't drop immediately), unlike typical deliquescent (rapidly disintegrating) spathe limbs in some other species.
Female zone bears about 120 pistils, ovary pale green, unilocular, apically incompletely 3-septate, style short, stigma bluntly lobed
Mature infructescences can have nearly full fertilization (over 98%), but unfertilized florets (small, seedless, orange, later reddish structures) often occur around fertile berries. Some unfertilized florets adjacent to berries may enlarge due to stimulation from a growth substance released by the developing berries, although remaining seedless.
POLLEN MORPHOLOGY:
The pollen of Alocasia brisbanensis is dimorphic (two forms):
Normal pollen grains: Larger (32.3–47.5 μm, mean 40.5 μm) with starch, viable.
Aborted pollen grains: Smaller (19.0–36.1 μm, mean 26.1 μm), without starch, inviable.
The presence of aborted grains is independent of genetic factors causing leaf variegation, indicating complexity in genetic expression
INFRUCTESCENCE:
Ripe fruits are bright red berries that split open longitudinally upon maturity. Seeds are brown to black, measuring about 3–4 mm in diameter, somewhat beaked, and strophiolate (having a small appendage at the base)
Fruit of Alocasia brisbanensis is ingested by various animal species, notably:
Eastern Water Dragon (Physignathus lesueurii lesueurii), observed consuming ripe red berries directly from the infructescence.
Birds including Regent Bowerbirds, Lewin’s Honeyeaters, and Brush-turkeys have also been recorded consuming its berries.
The fate of seeds after ingestion by these animals remains uncertain, suggesting an ecological research opportunity for Australian botanists
VARIEGATED FORMS: N/A
ETYMOLOGY: The specific epithet refers to the locality of Brisbane, capital of Queensland, in Australia
NOTES:
Hay (in press) has observed that, hitherto, the Australian representative of Alocasia has been repeatedly only partly described and appears to have been assumed to be the widespread and cultivated Alocasia macrorrhizos: an assumption based on superficial similarities. There are, however, numerous vegetative and reproductive differences, and there can be no doubt that this species is a very distinct Australian endemic.
Bailey (I.e.) intended his var. brisbanensis to be distinguished from most other Australian specimens on the basis of its smaller size and purple brown coloration in the petioles. Subsequently, Domin, who apperently did not really know the variety in detail, somewhat arbitrarily elevated it to specific rank. The differences that these authors intendedto highlight are, in view of comparison with variation in other Alocasia species, quite trivial. Here, the epithet ‘brisbanensis’ is used at specific level to denote all the known mainland Australian representatives. It does not seem useful formally to distinguish between the type kind and the supposedly larger green-leafed plants at any rank, unless horticultural distinction becomes desirable at the level of cultivar.
Hay defined Alocasia brisbanensis as belonging to a new, monotypic section called Ozarum, within Alocasia based on specific distinctive morphological characters:
Linear, often multistaminate synandria (pollen-producing units).
Robust and unusually long pedunculate inflorescences, which distinguish it from other Alocasia species
While obviously linear synandria are present in the male zone, some specimens show a preponderance of synandria with relatively few stamens, sometimes as few as two. However, even in cases where the numbers of stamens normal for most Alocasia species are found (4-6), their arrangement is quite different from that in other species. The anthers are arranged back to back, and the synandrial tissue consists almost entirely of anther, where in other species the anthers are arranged on or in the periphery of a block of connecting ground tissue.
The following character states distinguish Alocasia brisbanensis from Alocasia macrorrhizos:
ALOCASIA MACRORRHIZOS
i. Synandria isodiametric, stamens few
ii. Anthers opening by lateral slits
iii. Sterile interstice much shorter than female zone
iv. Inflorescence weakly scented
v. Limb of spathe cowl-like, membranous
vi. Peduncle shorter than inflorescence
vii. Posterior lobes usually rounded
viii. Glands present, conspicuous
ALOCASIA BRISBANENSIS
i. Synandria elongate, often multistaminate
ii. Anthers opening by apical pores
iii. Sterile interstice equalling to exceeding female zone
iv. Inflorescence powerfully fragrant
v. Limb of spathe spoon-shaped, rather leathery
vi. Peduncle much longer than inflorescence
vii. Posterior lobes of leaf usually acute to
rectangular
viii. Glands in axils of primary veins usually
inconspicuous or absent
4. Alocasia brisbanensis shows a distinct flowering peak, in December and January, throughout the range, though there are flowering and fruiting collections from all but the winter months. Shaw & Cantrell (1982) made a study of pollination and concluded that cross pollination by insects was indicated. A variety of insect taxa, including representatives of Diptera, Hymenoptera, and Coleoptera, were observed as visitors but none were conclusively demonstratedto be pollinators. They suggested, however, that Alocasia brisbanensis is most likely pollinated by Brachypeplus sp. (Coleoptera: Nitidulidae) or by an undetermined alaocharine staphilinid (Coleoptera). Shaw et al. (1985) noted fruits eaten by a Lewin's Honeyeater and a Regent Bowerbird in Queensland.
McAlpine (1978) described Neurochaeta inversa, a new species and genus representing a new family (Neurochaetidae) of Diptera, from the spathal chamber of wild Alocasia brisbanensis, citing it as ‘Alocasia macrorrhiza’.
Other specimens seen:
AUSTRALIA. Queensland, Moreton Dist., Brisbane, 3 mi Brush, Bailey 104 (BRI); Cook Dist., Julatten, Bales 158 (BRI); N.S.W., Hastings R., Beckler s.n. (MEL); Gloucester, Belche NSW 18914 (NSW); Queensland, Mt Glorious, Blake 12687 (NSW); N.S.W., Brickmaker's Ck, 4 mi E of Ramornie, Blakely <£ Shiress NSW 18913 (NSW); Georges Ck, NW of Kempsey, Boorman NSW 18917 (NSW); Queensland, Clarke Range, S Kennedy Dist., W of Cathu, Byrnes & Clarkson 3829 (BRI); N.S.W., Rivertree, Cambage 2872 (NSW, SYD); Byron Bay-Wingham (sic), Cleland s.n. (AD); Byron Bay, Cleland s.n. (AD); Queensland, Dalrymple Heights, Clemens s.n. (A); N.S.W., Breakfast Ck, foot of Mt Warning, 10 mi SW of Murwillumbah, Constable 3059 (NSW); Oaks State Forest, 20 mi SW of Bellingen, Constable 4818 (NSW); Ellcnborough R., 20 mi NW of Taree, Constable 6332 (NSW); back to Macquarie Falls, c. 12 km from Albion Park, Coveney 4825 (NSW); Whian Whian State Forest Reserve, 4-8 km SW of Dunoon, Coveney & Hind 10598 (NSW); Queensland, Wright Ck, Lake Eachan State Park, 20 km E of Atherton, Croat 52573 (NSW); Herbert R., Dallachy s.n. (MEL); Queensland, cult, at Forest Research Regional Station, Atherton, Dockrill 1100 (QRS); N.S.W., Acacia Ck, via Killarney, Dunn 272 (NSW); Queensland, Spicer's Gap, Everist 7286 (BRI, K); Mt Elliot, Fitzalan s.n. (MEL); N.S.W., Richmond R., Fitzgerald 1891 (MEL); Queensland, Russell R., Flecker 9678 (QRS); Wright's Ck, Flecker 9815 (QRS); N.S.W., Coffs Harbour, Floyd NSW 80709 (NSW); Mt Colah, Gillan NSW 18907 (NSW); Allyn R., Gordon NSW557097 (NSW); cult. University of Sydney ex Wauchope, Hay 4070 (NSW); Eastern Dorrigo, Hayes NSW 52512 (NSW); Queensland, Mt Tambonne, e. 45 mi S of Brisbane, Hubbard 2461 (L); State Forest Reserve 144, Agapetes L.A., Hyland 10181 (QRS); Timber Reserve 176, Monkhouse, Hyland 12194 (QRS); N.S.W., Yessabah Caves, Kempsey Dist., Ingram NSW 49352 (NSW); Queensland, East Barron, State Forest Reserve 191, Irvine 95 (BRI, QRS); Weinert's Ck, Bellenden Ker Range, Jago 214 (QRS); N.S.W., Kiama to Jamberoo, Johnson NSW 23705 (NSW); nr Big Hill, S of Crescent Head, Johnson NSW 91173 (NSW); Whian Whian, Jones 1239 (CANB); Queensland, Moreton Dist., Spicer's Gap, Kerr s.n. (BRI); N.S.W., Allyn Valley, Lawson s.n. (CANB, SYD); Queensland, 'Archer's', Leickhardt NSW 18908 (NSW); Queensland, Brisbane, Longman s.n. (K); N.S.W., Tooloom Range, Maiden NSW 18918 (NSW); Byron Bay, Maiden & Boorman NSW 18909 (NSW); Queensland, N Kennedy Dist., Kelsey Ck, nr Proserpine, Michael 1127 (BRI); 20 km SE of Atherton, Morawetz et al. 7-17185 (QRS); N.S.W., Table Top, Albury Dist., NSW 91104 (NSW); Manning R„ NSW 18910 (NSW); Hastings R., Oldfield s.n. (K); Queensland, Endeavour R., Persietz 1881 (MEL); Mt Glorious, Phillips 7829 (BRI); N.S.W., Bulladelah, Rupp s.n. (MEL); Banks of Dingo Ck, c. 5 mi NW of Wingham, Salasso 2837 (NSW); Clouds Ck, Salasso 4668 (NSW); Queensland, cult. CSIRO Longpocket, Brisbane, Shaw s.n. (BRI); Mt Glorious, Shaw s.n. (BRI); Peechey's Scrub, Simmonds 569 (BRI); Brookfield (Brisbane), Simmonds s.n. (BRI); Yandina, Simmonds s.n. (BRI);N.S.W., Pappinbarra, Simpson s.n. (SYD); Queensland, Mt Glorious, Smith 10542 (BRI); Darling Downs, c. 18 mi from Killarney, Head road, near the top, Stanley 7817 (BRI); N.S.W., Tweed R., Steel s.n. (BRI); Queensland, Rockhampton, Thozet s.n. (K); Theresa Ck, Millaa Millaa, Cook Dist., Trapnell 256 (BRI); cult. Brisbane Botanic Garden ex Indooroopilly, Trapnell s.n. (BRI); Ipswich, Vernsh 23 (MEL); N.S.W., Dorrigo National Park, c. 2 km down hill from the Glade, Wallace & Abel 103186 ; Queensland, end of Davies Ck road, Webb & Tracey 5595 (BRI); headwaters of Dryander Ck, Mt Dryander, Webb & Tracey 10101 (BRI); Cronin's Ck, between Mts Barney & Ernest, White 8761 (A, BRI); Mt Glorious, White 12079 (A, BRI); Queensland, Burke Dist., Croydon, Wilson 7 (BRI).
NEW ZEALAND. Along Route 5 between Tapapa and Rotorua, 35 km W of Rotorua, Croat 52233A (US)
CULTIVARS: N/A
HYBRIDS:
Alocasia 'Brisbane Blue' (Alocasia brisbanensis x Alocasia odora 'Blue')
Alocasia ‘Brisbane Tigress’ (Alocasia brisbanensis x Alocasia zebrina ‘Tigrina’)
Alocasia ‘Brisbane Waves’ (Alocasia brisbanensis x Alocasia portei)
Alocasia 'Tiny Dancers' (Alocasia brisbanensis × Alocasia odora)
















REFERENCES: